В соционике уже длительное время сосуществуют два разных (и, на первый взгляд, противоречащих друг другу) представления о формировании ТИМа. Согласно первому из них, ТИМ врожден и формируется генетическими факторами; к этому взгляду склоняется Е.С.Филатова [11, 12]. Согласно второму подходу, ТИМ формируется за счет раннего взаимодействия ребенка с окружающим миром (прежде всего, с матерью) либо за счет пренатальных влияний материнского организма на плод. Среди социоников, придерживающихся этого подхода – Е.В.Олькова; согласно ее теории, в ТИМе ребенка отражается характер проблем и надежд, переживаемых матерью во время беременности [5].
Как Филатова, так и Олькова исходят из собранной ими статистики типирования большого количества детей и родителей. Но, судя по всему, критерии типирования у них разнятся, из-за чего в их данных исходно содержатся разные закономерности. Филатова делает акцент на физическом (физиогномическом) сходстве людей одного и того же ТИМа и подтипа, после чего ссылается на мнение биологов, согласно которому "двойники" (люди, сильно похожие друг на друга) обладают очень близким генотипом (в т.ч. их ткани совместимы друг с другом при трансплантации) [12]. Но в то же время другие соционики наблюдали пары монозиготных близнецов (геном которых вообще идентичен) разного ТИМа. Сама Филатова считает, что МЗ близнецы должны иметь одинаковый ТИМ, но опирается не на своих данных, а на наблюдениях другого соционика – Д.А.Иванова.
В более широком плане, наследование психологических качеств и черт характера составляет предмет психогенетики – науки, основанной в XIX столетии Ф.Гальтоном. В психогенетике накоплены данные о наследовании экстра-/интроверсии по опросникам Кеттела и MMPI (типологией Юнга психогенетика пока не занималась). Обнаружена высокая наследуемость – корреляция порядка 50% для МЗ близнецов и 20% для обычных братьев/сестер [3, §11.2], но полного совпадания вертности у МЗ близнецов нет.
Но психогенетика исследует и так называемые материнские эффекты – внутриутробное либо постнатальное влияние матери на психические качества, проявляющиеся в потомстве уже в зрелом возрасте. Существование таких эффектов доказано, в т.ч. строго контролируемыми экспериментами на животных. Так, в опытах на мышах было показано, что постнатальные материнские влияния способны снижать агрессивность самцов, происходящих из генетически более агрессивной линии. Другими словами, материнский эффект может усиливать либо подавлять действие обычных генов; в итоге на один и тот же признак могут совместно влиять гены, внутриутробные влияния во время беременности, и особенности ухода за потомством [3, §9.6].
Кстати, косвенное подтверждение наличия материнского эффекта в наследовании ТИМа можно найти в семейной статистике Е.С. Филатовой [11, 6] (хотя сама Филатова придерживается мнения о полностью генетической природе ТИМа). Эффект можно увидеть, если разделить семьи, входящие в статистике Филатовой, по признаку экстра-/интроверсии на две группы: в первую группу – семьи, в которых мать и отец различаются по признаку вертности, во вторую – если родители имеют одинаковую вертность. Что показывает статистика наследования этого признака? В первой группе дети чаще наследуют вертность отца (с вероятностью выше 2/3) и реже – матери; однако во второй группе вертность детей примерно в половину случаев противоположна вертности обоих родителей! Это очень сложно объяснить одной лишь классической генетикой. Согласно последней, если проявленность (фенотип) признака у обоих родителей совпадает, дети с противоположным фенотипом признака хотя и рождаются (скажем, унаследовав рецессивные гены), но это происходит никак уж в половину случаев, а гораздо реже. "Нормальная" (в смысле менделевской генетики) картина наследования в статистике Филатовой наблюдается, что примечательно, только в отношении признака рацио-/иррациональность. Ряд других признаков (вертность, статика-динамика, часть признаков Рейнина) ведет себя описанным выше образом, когда дети как будто "отталкиваются" от матери по этим признакам. Общий результат по всей статистике и всем признакам выражается в повышенном числе детей, дуальных и противоположных матери.
Это отталкивание можно назвать обратным материнским эффектом (ОМЭ). Е.Олькова тоже упоминает похожую закономерность: у экстравертных матерей рождаются больше интровертных детей, и наоборот (хотя Олькова объясняет этот эффект совершенно по-другому). Часть вероятных механизмов ОМЭ будет обсуждена ниже; сначала сделаем еще пару важных замечаний.
Первое из них касается распределения соционических признаков в крупной популяции (типа этноса). Если бы признаки определялись действием одних только генов в узком смысле, было бы возможно путем отбора сформировать популяции, в которых соотношение носителей разных признаков (скажем, экстра- и интровертов) различалось бы очень сильно, на порядок. Но такая популяция была бы очень несбалансированной в плане социальных взаимодействий; ведь ТИМ – это, кроме прочего, базисная социальная роль (на уровне этологии). В.В.Гуленко показал, что при взаимодействии тождественных ТИМов они "отталкиваются" друг от друга, занимая в группе разные роли в зависимости от подтипов [1]. Но с точки зрения эффективности лучшее решение – сбалансированность по признакам, когда на роли экстра- и интровертов, рацио- и иррационалов и т.д. претендует примерно одинаковое количество одинаково подходящих (в смысле ТИМов) людей; в противном случае даже если роли худо-бедно распределены, возникнет нескомпенсированный избыток "лишнего" социального действия по одним признакам.
Вот здесь и оказался бы крайне полезным обратный материнский эффект. Ведь он постоянно превращал бы часть "генетических экстравертов" в соционические интроверты (и наоборот), компенсируя таким образом избыток людей одного полюса в сторону другого. Разумеется, в масштабе популяции эффект действует лишь в среднем, в статистике; признаки же каждого отдельного человека зависят только от его родителей.
Такие признаки будут состоять из двух компонентов: генетический и эпигенетический, т.е. обусловленный материнским эффектом. При этом среднепопуляционное значение первой составляющей может быть сдвинуто в ту или иную сторону, что, скорее всего, обуславливает интегральный ТИМ популяции (этноса, субэтноса). Этносы в самом деле отличаются, прежде всего, средней яркостью (в основном – невербальных проявлений) соционических признаков. Так, темперамент северных и южных европейцев в среднем заведомо отличается, но значит ли это, что среди северян численно преобладают (притом в значительной мере) соционические интроверты, а среди южан – соционические же экстраверты? Отнюдь. Ведь часть "генетических интровертов" материнский эффект вытолкнул в соционические экстраверты, и наоборот. Притом в смешанных группах со стороны вполне может казаться, что признак зависит в большей степени от генетического фактора, чем от эпигенетического (на нашем примере экстраверты-северяне будут казаться интровертнее интровертов-южан). К вопросу о разделении и опознавании генетических и эпигенетических факторов в поведении человека мы вернемся еще раз.
Второе замечание касается так называемого хэндлинг-эффекта [3, §9.6; 13]. Он обнаружен при изучении влияния раннего опыта на поведение животных. Оказалось, что если детенышей мышей и крыс в течение первых трех недель после рождения регулярно берут в руки (англ. handling), это оказывает долговременное влияние на их поведение (по сравнению с контрольными животными, не получавшими хэндлинга, даже если последний ограничивался тремя минутами в день). Став взрослыми, особи, подвергавшиеся детскому хэндлингу, демонстрируют пониженную эмоциональную реактивность, низкую чувствительность к стрессу новизны, а также – как следствие последнего – большую любознательность. Показано, что хэндлинг уменьшает напряжение гипоталамо-гипофизарно-надпочечниковой оси и повышает адаптационные способности на стресс.
Изначально считалось, что изменение поведения животных – прямое следствие хэндлинга. Однако последующие наблюдения за взаимоотношениями животных, подвергавшихся хэндлингу, с их матерями показало, что после хэндлинга и у мышей, и у крыс матери гораздо больше "тормошили" и вылизывали своих детенышей (раздражителем был запах человека, который мать старалась устранить), гораздо чаще принимали позу "нависания" над детенышем и т.д. Именно эта повышенная стимуляция со стороны матери приводит к стойкому снижению эмоциональности у потомства. Если же детеныши матери с высокой активностью выращиваются приемными матерями с низкой активностью, их эмоциональность повышается. Это доказывает, что главное значение имеет эффект раннего опыта.
Последствия хэндлинга с точки зрения соционики выглядят как подавление
ЧЭ и ЧС (гипоталамо-надпочечниковая ось связана с нейромедиаторами
норадреналин и адреналин), с некоторым сдвигом к интуиции; причину же их
возникновения лучше интерпретировать, как усиление ЧС-действия со стороны
матери. Но дело не в точном "типировании" эффекта, а в самом свидетельстве
наличия такого механизма регуляции поведения у животных.
Вернемся к вопросу формирования ТИМа. Мы уже показали в других работах
[7, 8], что все признаки Юнга являются по сути двухкомпонентными.
Во-первых, есть активационная (энергетическая) составляющая, отражающая
активность разных систем мозга (в основном, подкорковых). И во-вторых, есть
собственно информационная составляющая ТИМа, отражающая представленность
юнговских функций в исполнительной системе мозга (префронтальные доли коры)
за счет преимущественной связности последней с разными участками коры и с
разными активационными системами подкорки. В первом приближении можно
принять, что энергетическая составляющая формируется за счет мощности
определенных систем мозга, а информационная – за счет мощности связей
между ними (очень схематическое представление этой картины показано на
фиг.1).
Но если механизмы действия двух составляющих признаков разные, то
механизмы их формирования тоже могут существенно отличаться. Поскольку мы
здесь обсуждаем соотношение генетических и эпигенетических (обратный
материнский эффект) факторов, поставим следующий вопрос: могут ли эти
факторы действовать по-разному на информационную и на энергетическую
составляющую признаков? В качестве предельного случая – может ли одна из
составляющих формироваться полностью за счет генетического фактора, а
вторая – за счет ОМЭ?
Эксперименты на животных (в т.ч. с хэндлингом) здесь малоинформативны,
т.к. в поведении животных выявляется по сути только энергетическая
составляющая. В ней, как видно из приведенных примеров, могут вносить вклад
оба механизма. Но оценить связностную составляющую таким образом вряд ли
возможно, если вдобавок иметь в виду, что у животных префронтальная кора
гораздо меньше по относительному размеру, чем у человека, и ее вклад в
поведении не в пример слабее.
Очень ценными и значимыми здесь могут оказаться данные близнецовых
исследований. В них изучается наследование одних и тех же признаков у
монозиготных (однояйцевых) и дизиготных близнецов, имея в виду, что первые
являются генетически тождественными, а вторые – нет (с т.з.
генетики они суть обычные братья и сестры). Исследования утверждают, что МЗ
близнецы очень схожи по признакам темперамента: для признаков вроде
активность/пассивность, устойчивость внимания, порог реактивности и проч.
получаются внутрипарные корреляции до 90% и выше [3, §11.2], что по меркам
психогенетики очень много. (Притом те же корреляции у дизиготных близнецов
зачастую оказываются очень низкими и даже отрицательными, что совершенно не
характерно для наследования количественных признаков.) Для более "сложных"
факторов личности, выявляемых по опросникам MMPI, Кеттела и другие,
внутрипарные корреляции у МЗ близнецов заведомо ниже и редко превышают 60%.
На первый взгляд сравнение МЗ близнецов вообще ничего не может сказать
о действии материнских эффектов, т.к. последние действуют одинаково на обоих
близнецов и накладываются на их одинаковый же генотип. Однако, кроме
материнских эффектов, здесь мы имеем такие же воздействия близнецов друг на
друга, которые по механизму могут вообще не отличаться от воздействия
матери. В частности, если близнецы растут вместе, они постоянно общаются
еще с самого рождения, – разумеется, сначала лишь невербальными сигналами,
но как раз они (по нашей гипотезе) и являются решающими для обратного
материнского эффекта. Результат этого взаимодействия можно назвать
обратным близнецовым эффектом (ОБЭ); он должен приводить к расхождению
некоторых поведенческих (в т.ч. соционических) признаков, в результате чего
даже МЗ близнецы будут демонстрировать разный ТИМ.
В принципе, более высокую корреляцию "энергетических", темпераментных
факторов по сравнению с более "высшими" свойствами личности в паре МЗ
близнецов можно объяснить более сильным влиянием социальной среды на
"высшие" свойства. Но исследования также показали, что у МЗ близнецов
отличаются и другие фундаментальные психофизиологические признаки, в
частности – функциональная асимметрия мозга. Так, у них разная
асимметрия фоновой ЭЭГ [9]: если один близнец имеет выраженную
латеральную асимметрию ЭЭГ, то у второго, как правило, картина ЭЭГ
симметрична (отсутствуют собственно зеркальные пары, т.е. МЗ близнецы с
выраженной асимметрией противоположного знака). МЗ близнецы могут
отличаться "рукостью" – один может быль левшей, второй – правшей, с
вероятностью более 20% (притом для ДЗ близнецов вероятность примерно та
же).
Итак, имеем разницу в асимметрии ЭЭГ МЗ близнецов при значительно
более одинаковом уровне темпераментных признаков. Но последние отражают
активность центров подкорки, асимметрия же ЭЭГ формируется вследствии
неодинаковой активности разных зон коры (а также – неодинаковой их
активации одними и теми же центрами). Это значит, что у МЗ близнецов
есть существенные отличия в мощности связей, по которым активация
передается к коре. Формируются же эти отличия эпигенетическими факторами
(раз геном идентичен), в т.ч., возможен описанный выше обратный эффект.
Очень важые результаты получены А.А.Ивониным с соавторами [2]. В
близнецовом исследовании они установили, что наиболее жестко генетически
детерминируется развитие подкорковых структур среднего мозга и ствола
мозга, а также ассоциативных таламо-кортикальных систем внутримозговой
интеграции. В развитии межрегиональных связей новой коры наследственные и
средовые факторы действуют неоднозначно, притом формирование длинных
путей обусловлено в большей мере генотипом, а более близких межцентральных
связей в пределах каждого из полушарий – в большей мере влиянием внешней
среды.
В качестве итога можно выдвинуть следующее общее положение:
К этому результату можно прийти и с другой стартовой точки, взяв за
основу факт, что подкорковые центры эволюционно древнее, а исполнительные
центры новой коры, связность которых со специфическими отделами мозга
определяет ТИМ, – намного моложе (они ведь охватывают самую молодую,
третичную ассоциативную кору, интенсивно развившуюся лишь у крупных
приматов). Но более молодые структуры вместе с тем более пластичны в
онтогенезе, сильнее реагируют на изменения среды (а материнский эффект –
суть влияние среды, только более узкого плана).
Будем полагать, что эти сигналы, являясь врожденным этологическим
базисом раннего общения, распознаются разными системами мозга младенца,
которые примерно соответствуют соционическим функциям. Далее, в ответ на
сигналы формируется реакция младенца, в т.ч. ответные сигналы (здесь
работают те же системы-функции мозга).
Также, интенсивность и модальность внешних сигналов сказываются на
развитие функций следующим образом: чем избыточнее и интенсивнее внешние
сигналы по данному аспекту, тем более развитие данной функции подавляется.
В частности, сам ребенок в своих ответных сигналах начинает избегать этот
аспект; при этом может расти интенсивность других функций, которых младенец
и будет предпочитать для своих ответных сигналов.
Подавление функции, как мы предположили, должно происходить за счет
относительного ослабления каких-нибудь связей в мозгу, – точнее, не
собственно ослабления, а подавления их развития в критическом периоде
онтогенеза (хотя конечный результат тот же самый). Допустим, со стороны
матери избыточны сигналы по ЧС. Это должно подавить, прежде всего,
иррационально-экстравертный темперамент, точнее, связи соответствующей
подкорковой системы с исполнительной системой мозга (префронтальная кора).
На фиг.2. связи подкорковых темпераментных систем с ПФК даны внизу, а
подавление роста связей обозначено тонкими красными линиями. Связи других,
неизбыточно представленных во внешних сигналах темпераментов с ПФК будут
развиваться нормально и представительство других темпераментов в ПФК
возрастет. Один из них (скорее всего, сильнейший по генетическому фактору)
станет доминировать в поведении – даже если "его" подкорковая
активационная система объективно слабее ЧС-системы (на фиг.2 мощность
активационных центров обозначена синими столбиками снизу). Это очень важное
положение.
Должны ли ослабнуть также связи ПФК с сенсорными отделами коры (на
фиг.2 – верхний ряд связей), придающие аспекту ЧС собственно многомерность?
Здесь ситуация сложнее. Прежде всего сигналы, которыми обмениваются мать и
ребенок, в целом просты и инстинктивно понятны, для их понимания и
обработки высшие размерности функции вообще не нужны. Да и на столь ранней
фазе развития мозг вряд ли способен обеспечить эти высшие мерности.
Однако здесь может иметь место другой эффект. В [8] мы уже писали, что
кроме "многомерных" исполнительных центров в префронтальной коре есть также
дополнительные "маломерные" (по критериям соционики) центры, локализованные
в теменно-височных областях. Они берут на себя интеграцию простых либо
давно знакомых, отработанных до автоматизма действий, в которых уже можно
обойтись без вмешательства мощной, но энергоемкой ПФК. Так вот, избыточность
сигналов по одному из аспектов (скажем, ЧС) со стороны матери приведет к
тому, что ребенок очень быстро привыкнет к ним, вслед за чем мозг "спихнет"
опознавание и реакцию на них менее мощным, но менее энергоемким маломерным
центрам. В итоге станут быстрее развиваться связи маломерных центров со
специфическими областями мозга, в которых осуществляется первичная обработка
данного аспекта (в нашем случае, сенсорика). И наоборот, нехватка внешних
сигналов по противоположному аспекту (в случае, интуиции) приведет к тому,
что младенец медленнее привыкнет к ним, мозг продолжит обрабатывать ее
"многомерной" ПФК; таким образом связи специфических областей коры с ПФК
усиливаются, связи же с "маломерными" исполнительными центрами развиваются
слабее. Связи двух исполнительных центров показаны на фиг.3. (Отметим, что
информационное воздействие идет "сверху вниз", от специфических отделов
коры к ПФК; активация же передается снизу вверх, от подкорковых центров к
ПФК).
Итак, мы описали два возможных эпигенетических механизма обратного
материнского эффекта при формировании признаков: а) путем ослабления (или,
точнее, недоразвития) связи ПФК с одним из центров активации, и б) путем
переключения обработки информации от многомерного (ПФК) к маломерному
(теменно-височному) исполнительному центру. Но мы допускаем также
существование других похожих механизмов, отличающихся в деталях от
описанных здесь. Еще раз, цель этого текста – показать в наиболее общем,
схематичном виде принципиальную возможность взаимодействия генетических и
эпигенетических факторов ТИМа, на уровне активационных функциональных
систем и их связей с исполнительными центрами мозга.
Приступим к анализу этого взаимодействия. По конкретному признаку
генетические и эпигенетические факторы могут действовать в одну сторону
либо, наоборот, "тянуть" признак в разные стороны. В первом случае картина
проста и предсказуема: к примеру, ОМЭ подавляет связи с "изначально"
слабым центром активации, в итоге развиваются связи с сильнейшем центром и
он становится абсолютно доминирующим. Гораздо интереснее второй случай, в
котором имеет место противоречие, конфликт действующих механизмов двух
родов. Рассмотрим поподробнее эти противоречия и способы их разрешения в
онтогенезе.
Допустим, что "под гнетом" ОМЭ попал сильнейший подкорковый центр и
его связи с ПФК подавляются. Это может привести к некоторому подавлению
активности самого центра, но он все равно останется сильным. В то же время
его активация не востребована ПФ корой. Куда она денется? Во-первых, ее
"истратят" специфические центры, не связанные непосредственно с мышлением,
– скажем, подкорковые двигательные центры (что отразится в общей активности
человека). Во-вторых, что гораздо важнее, избыток активации может
направиться в маломерные исполнительные центры. Они относительно
независимы от многомерных (в лице ПФК) и заняты рутинными действиями-
автоматизмами, в которых можно обойтись без высших размерностей функций и
блоков [8]. В итоге окажется, что по маломерным функциям человек работает
"не в том темпераменте", как по многомерным: скажем, ИЛЭ при решении
этических либо сенсорных задач может вести себя скорее как интроверт,
и/или как рационал. Кстати, по данным В.Л.Таланова [10], это нередко и
происходит (хотя Таланов интерпретирует этот факт в рамках своей модели Т).
В.В.Миронов также предлагал схему распределения темпераментов по блокам
модели А [4].
Итак, противоречие факторов двух родов выливается в том, что в работе
многомерного исполнительного центра доминирует не сильнейший (по абсолютной
мощности) темперамент, а другой (вероятнее всего – второй по силе); в то же
время первый может доминировать в маломерном центре (фиг.4). Это –
разрешение противоречия в виде разделения в пространстве: в ТРИЗ
(теории решения изобретательских задач) он является одним из универсальных
методов разрешения противоречий, наряду с разделением во времени (когда
конкурирующиеся центры чередуются в доминировании). Последнее тоже имеет
место в психике человека, ведь темпераментные центры способны менять свою
активность согласно требованиям ситуации. Однако нас здесь интересуют
консервативные, не зависящие от текущей ситуации свойства соционических
функций: согласно [7], они определяются системой дальних связей в мозгу,
последние же относительно стабильны, поэтому разделение во времени для них
не работает.
Разделение в пространстве, как способ разрешения конкуренции разных
систем мозга, может проявляться и другими способами. Вспомним, что
исполнительных центров на самом деле четыре [8]: в лобной и теменно-
затылочной коре, слева и справа. Два из них многомерны, два – маломерны,
притом они относительно независимы друг от друга. Как результат, один из
многомерных центров может быть преимущественно связан с одним темпераментом
и с одним блоком функций, второй – с другими (фиг.5). Из двух этих центров,
судя по всему, определяющим для ТИМа является тот, который находится в
речевом полушарии: ведь ТИМ, по мнению подавляющего большинства социоников,
отражается в лексике и в структуре речи. (Как правило, речевое полушарие у
правшей – левое, у левшей же бывает по-разному). Однако второй, неречевой
исполнительный центр может задавать подтипное усиление, в виде повышенной
склонности к обработке информации в данном режиме, но без должного ее
прорабатывания в речевом мышлении и, как следствие, без достаточной
осознанности процесса.
Разделение функций по полушариям может разрешить возможный конфликт
двух механизмов роста связей (хотя мы считаем, что связи "программируются"
в основном эпигенетикой, на них все-таки влияют и генные факторы, см выше).
Допустим, что генными факторами задана сильная связь исполнительных центров
с сенсорикой, а ОМЭ, наоборот, "выдавливает" сенсорику в маломерные центры.
Однако он может ее выдавить только из ПФК ведущего полушария, оставив сильную
ее связь с ПФК противоположного полушария (фиг.5). Таким же образом в принципе
возможна и латерализация связей ПФК с темпераментными центрами активации:
левая ПФК "замкнута" в основном на один темперамент, правая – на другой.
В этом ключе очень показательны результаты исследования Т.А. Мешковой
асимметрии биоэлектрической активности коры (ЭЭГ и вызванные потенциалы) у
близнецов (в изложении [9]):
Что это значит? Напомним, левое полушарие в норме специализировано на
работу с социальными сигналами (не только речь и не только у человека, но и
у других видов, в т.ч. оно отвечает за песню птиц). Преобладание средовых
факторов в развитии левого полушария означает, что действуют в основном
социальные сигналы – как раз по типу предположенных нами эпигенетических
эффектов (в т.ч. близнецовый). В противоположном же полушарии остается
действие преимущественно генных факторов. Так что наша гипотетическая модель
формирования ТИМа вполне согласуется с результатами Т.А. Мешковой. Замечание
же о том, что асимметрия четче всего обнаруживается в височных областях,
возможно, указывает на преимущественную роль не префронтальных (многомерных),
а наоборот, височно-теменных (маломерных) исполнительных центров в
формировании ТИМа. В самом деле, в этом возрасте ПФ кора еще не сформирована,
да и согласно нашей модели сначала развиваются связи с маломерными ИЦ и уже
потом – с многомерными (см. рис.3).
(Еще одно замечание. Совместимость нашей модели с картиной
функциональной асимметрии активности коры у близнецов указывает на то, что
лучшим критерием определения ТИМа, скорее всего, должна стать асимметрия
ЭЭГ и ВП типируемого – фоновая либо при решении разных задач. Но это скорее
относится к задачам будущего развития соционики.)
Наконец, принцип разделения в пространстве для разрешения противоречий
может действовать и внутри отдельно взятого исполнительного центра. Ведь
последний поддерживает связь со всеми функциональными отделами, только одни
связи мощнее, другие – слабее, за счет чего центру доступно избыточное
количество ресурсов (информация, активация) с первых, и недостаточное – с
вторых. Разделение в пространстве может вылиться в том, что разные функции
и блоки "скрещены" преимущественно с разными темпераментами. Допустим, что
область ИЦ, на которую замыкаются сильнейшие связи из, скажем, сенсорно-
логических отделов, "подпитываются" в основном рационально-интровертной
активацией. Но в другой, меньшей по размеру области ИЦ, поддерживающей
связь с интуитивно-логическими отделами, может доминировать другой тип
активации, скажем, иррационально-интровертный (фиг.6).
Разумеется, в разных исполнительных центрах функции и блоки могут
быть "скрещены" с темпераментами по-разному. Пример: левая ПФК замкнута
преимущественно на сенсорику-логику и рацио-интроверсию (ЛСИ), правая – на
логику-интуицию и иррац-экстраверсию (ИЛЭ). В таком случае в поведении будет
наблюдаться относительное уравновешивание признаков сенсорика/интуиция,
вертность и нальность. Но вместе с тем будут усилены признаки Рейнина,
образованные на пересечении ослабленных признаков Юнга: в случае –
"тактика" и "беспечность" (поскольку в мышлении сенсорика будет завязана на
интроверсию-рациональность, интуиция – на экстраверсию и иррациональность).
Здесь мы видим, что выраженность признаков Рейнина отнюдь не обязательно
"складывается" из выраженности образующих признаков Юнга: наоборот,
возможны случаи, когда ПР, образованный двумя относительно размытыми
признаками Юнга, выражен очень ярко за счет усиленной асимметрии
"скрещивания" информационных и активационных связей. Этому взгляду, кстати,
придерживается ряд социоников – В. Миронов, А. Трехов, А. Букалов.
Итак, мы показали, что конкуренция генетических и эпигенетических
факторов формирования ТИМа в онтогенезе разрешается вполне "мирными" и
стандартными способами. Более того, психике, сформированной в условиях
сильной конкуренции факторов, потенциально доступны более разнообразные
режимы информационного метаболизма. Притом последний оказывается заведомо
шире рамок модели "А" и теории Рейнина. Исходя из модели распределения
активности и функциональных связей по исполнительным центрам, в принципе
можно построить новую, более сложную теорию подтипов, учитывающую разные
структуры взаимодействия активационных и информационных факторов ТИМа (=
активационных центров и связей с префронтальными исполнительными центрами).
Одно из возможных решений показано в [8]. Однако окончательного решения
вопроса о подтипах у нас еще нет, потому что конкуренция факторов пока
проработана лишь в общем виде, без должной конкретики (нейрофизиологические
факторы информационного метаболизма пока установлены лишь в приближенном
виде). Да и методы типирования по поведению и речи пока не годятся для
определения большого количества факторов (мощность центров и связей всех
четырех исполнительных систем мозга). Возможно, здесь помогут средства ЭЭГ
и томографии.
Возвращаясь к вопросу взаимодействия факторов, можно подытожить, что
чем больше факторов в мозге (и психике) работают друг против друга,
конкурируясь, тем "гармоничнее" выглядит психика и личность. Разные мозговые
системы и связи дают сложную мозаичную картину активности, в которой
проявляются разные функции (но каждая из них – со своими особенностями;
скажем, одна из них многомерна, но замкнута на энергетически слабый
активационный центр, другая – наоборот, и пр.). Психика же, в которой
факторы не конкурируются, а складываются, действуя в одну сторону, заведомо
проще и беднее по своим проявлениям (хотя типируются такие люди намного
легче и однозначнее, чего очень сложно добиться в случае конкуренции многих
факторов). Она же имеет больше слабых мест и "недоступных" режимов работы,
хотя это часто компенсируется выдающимися способностями сильных – и даже
чересчур усиленных – функций и режимов работы.
Однако основные эпигенетические воздействия, как мы уже выяснили, –
обратные эффекты: по какому-то признаку ребенок "отталкивается" от того
же признака матери. Притом отталкивание идет за счет невербальной, базисно-
этологической и по сути энергетической стороны взаимодействия матери с
ребенком. Но "энергетику", как было уже сказано, определяют преимущественно
генетические факторы. Следовательно, фенотипический признак (скажем,
вертность) ребенка "отталкивается" от того же генотипического фактора
матери, – и складывается с генотипическим же фактором, унаследованным у
одного из родителей. Если у матери, они уравновешиваются описанными выше
механизмами конкуренции. Но если второй фактор унаследован у отца, и если он
сильно отличается от материнского фактора, они не погашаются, а наоборот,
усиливают друг друга.
Получается очень любопытное следствие: чем дальше друг от друга
отстоят генотипические признаки отца и матери, тем больше вероятность
появления потомства, сильно несбалансированного по данному признаку. К
примеру, самые "глубокие" интроверты будут рождаться не от двух
интровертных родителей, а от интровертного отца и экстравертной матери
(здесь "вертность" родителей – в смысле генного фактора). Эта картина
разительно отличается от обычного для психогенетики наследования
количественных признаков, но мы уже показали, что соционические признаки –
двухфакторные.
Отметим, что наибольшую вероятность для наибольшей разницы по генным
факторам – и, как следствие, высокую вероятность рождения сильно
несбалансированного потомства – дают межрасовые скрещивания. Далее следуют
родительские пары из разных этносов или субэтносов (последние иногда
заметно отличаются по темпераменту). Но в принципе эта вероятность
высока в любом случае, если родители сильно отличаются друг от друга по
темпераментным факторам. Еще раз подчеркнем: это вообще не вписывается в
классическую психогенетику количественных признаков. Однако похожий эффект
давно знаком селекционерам: при скрещивании далеких друг от друга пород
(скажем, собак) с высокой вероятностью рождается сильно неуравновешенное по
поведению потомство. Это – очередное свидетельство в пользу того, что
психические функции и ТИМ человека развиваются и функционируют по законам,
общим для большинства высших животных, а не являются чем-то уникальным для
человека (хотя, разумеется, человеческая психика намного сложнее животной).
1. Гуленко В.В.
Теория подтипов: система DCNH.
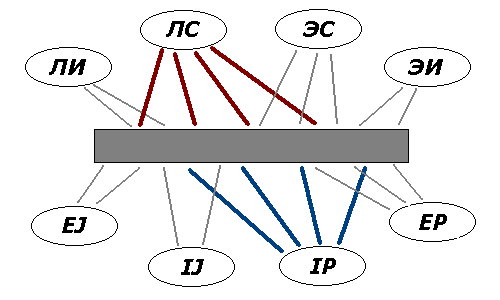
Фиг.1. Исполнительная система мозга (темный прямоугольник)
и ее связи с функциональными зонами коры (сверху) и с активационными
системами подкорки (снизу). Жирными линиями даны доминирующие связи.
Функциональные зоны коры, согласно [8], в первом приближении
соответствуют соционическим клубам, активационные центры – темпераментам.
Для признаков экстра/интроверсии и рацио-/иррациональности взяты
обозначения из MBTI (E/I, J/P).
Теперь перейдем к более конкретному описанию механизмов формирования
ТИМа на уровне нейрофизиологии. Будем считать, что эпигенетические
механизмы определяются сигналами (невербальными) со стороны ближайшего
окружения ребенка (в первую очередь – матери) в первых месяцев жизни
(максимум до 1.5 – 2 года). Сигналы – это и выражение лица, и взгляд, и
тон голоса, и движения рук, в т.ч. манипуляции с самим ребенком. Они
различаются по интенсивности, модальности, по преобладанию и избыточности
одних форм за счет нехватки других, – все это суть индивидуальный стиль
общения матери с ребенком, зависящий, кроме прочего, от соционических
признаков.
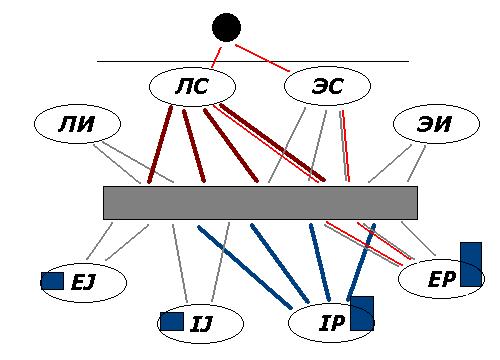
Фиг.2. Влияние средового фактора (избыток влияния по
аспекту ЧС) на развитие связей исполнительного центра со специфическими
системами мозга. Обозначения такие же, как на фиг.1; тонкой красной линией
показан путь ЧС-сигнала, темно-синими столбиками внизу – мощность
темпераментных активационных центров.
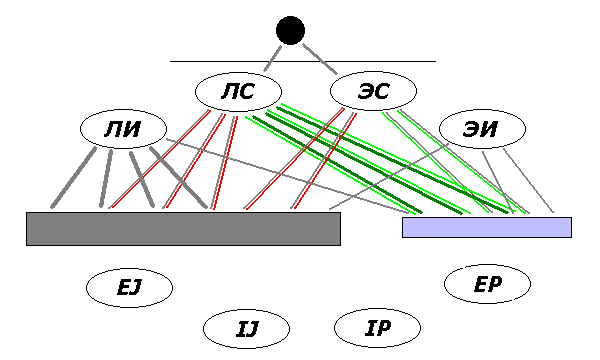
Фиг.3. Фактор среды (избыток влияния по ЧС) стимулирует
связи "своих" функциональных зон коры с маломерным исполнительным центром
(справа, тонкая зеленая линия) и подавляет их же связи с многомерным
центром (слева; тонкая красная линия). В итоге в многомерном центре
доминирует связь с одной из зон противоположной функции
(логико-интуитивная, см. жирные линии).
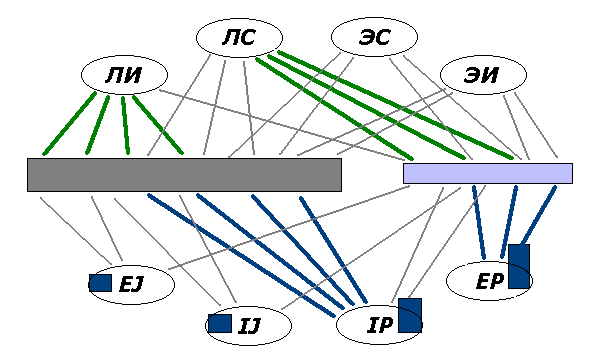
Фиг.4. В многомерном и маломерном исполнительных центрах
доминируют связи с разными специфическими системами (см. жирные линии),
из-за чего первый "сконфигурирован" как ИЛИ (и работает в таком режиме), а
второй – как СЛЭ.
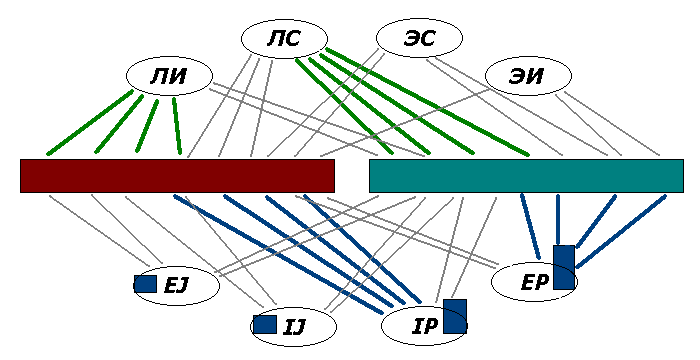
Фиг.5. В многомерных исполнительных центрах в левом и правом
полушарии доминируют разные связи; левый (доминантный по речевому мышлению)
"сконфигурирован", как ИЛИ, правый – как СЛЭ.
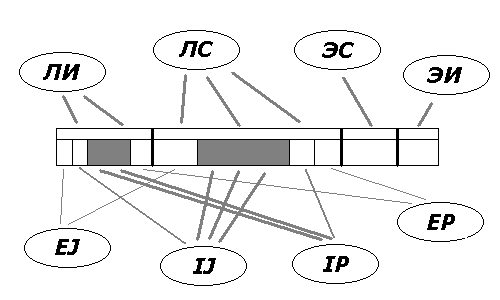
Фиг.6. Неоднородное скрещивание связей с разными функциональными
системами внутри одного и того же исполнительного центра. Доминируют
конфигурации типа ЛСИ и ИЛИ. Литература
2. Ивонин А.А., Цицерошин М.Н., Погосян А.А.,.Шуваев В.Т. Генетическая
обусловленность нейрофизиологических механизмов корково-подкорковой
интеграции биоэлектрической активности мозга. // Рос.физиол. журн.2002.
Т.88,№10. С.1330-1342. (тезисы здесь)
3. Мешкова Т.А., Николаева Н.О.
Психогенетика (Интернет-учебник, 2004).
4. Миронов В.В.
Несколько замечаний о модели А (2007).
5. Олькова Е.В. «Происхождение соционического типа, или проблемы в наследство» (2007)
6. Попов И.
О наследовании соционических признаков: Технический анализ статистики
Е.С.Филатовой (2006).
7. Попов И. Связи (блоки)
соционических функций (2010).
8. Попов И.
Нейрофизиологическая природа размерности функций (2011).
9. Равич-Щербо И.В., Т.М. Марютина, Е.Л. Григоренко. Психогенетика. //
Под редакцией И.В. Равич-Щербо. – М., 1999 //
с. 349-362.
10. Таланов В.Л.
Краткое изложение модели «Т»: физиологической модели информационного
метаболизма в психике человека
11. Филатова Е.С.
Соционическая статистика для 299 женщин, мужчин и их детей //
«Соционика, ментология и психология личности», Киев, 2000, № 6 (33).
12. Филатова Е.С.
Соционика и генетика // Филатова Е.С. Личность в зеркале соционики:
Разгадка тайны двойников. – СПб.: Б&К, 2001. – 286 с.
13. Фридман В.С.
Сигнальная наследственность в материнском поведении млекопитающих
Иван Попов, февраль 2011
![]()