Введение
Данным текстом я продолжаю ряд более ранних работ [9, 10, 11], посвященных нейрофизиологическому базису соционики. Однако прежде чем перейти к существу, придется сделать несколько общих теоретических замечаний.
Они касаются проблемы сводимости психических феноменов к конкретным нейрофизиологическим механизмам. В общем виде проблема такая: допустим, психологи выделили в поведении человека какие-то феномены (пример – работа соционических функций) и научились их опознавать. Но значит ли это, что каждому феномену должен соответствовать свой, особый нейрофизиологический механизм (либо система мозга)? Отнюдь. Ведь понятия и категории, в рамках которых мы описываем психику, нередко вводятся сугубо умозрительно, как отражение подвернувшихся авторам философских или бытово-психологических понятий. Разумеется, какие-то связи психического с нейрофизиологическим безусловно существуют, наука пусть и медленно, но вполне результативно продвигается в эту область познания. Но результаты, как правило, носят характер не прямых связей, а скорее статистических корреляций ("у 75% пациентов с повреждениями данной зоны коры выявлены такие-то нарушения"). Более того, в силу пластичности психики и слишком общего характера большинства тестовых задач, нередко одна и та же задача может решаться за счет действия совершенно разных систем мозга.
Для иллюстрации рассмотрим такую аналогию. Допустим, мы наблюдаем полеты самолетов разных типов и отмечаем, что они маневрируют и ведут себя по-разному. К примеру, видим наличие и количественное преобладание разных фигур пилотажа (бочка, иммельман, петля Нестерова и проч.), и на основе этого "типируем" самолеты. Так вот, каким образом связаны наши фигуры пилотажа с конструкцией самолета? Безусловно, какая-то связь есть: для некоторых фигур нужна большая тяговооруженность, для других маневренность, для третьих – устойчивость полета на больших углах атаки (либо, наоборот, динамическая неустойчивость в полете); некоторые требуют отклонение вектора тяги. Но как бы ни изучали и разбирали мы самолет, невозможно выделить в нем отдельные системы, каждая из них отвечающая в одиночку за иммельман, за петлю Нестерова и проч. Во всех фигурах задействованы одни и те же органы управления (аэродинамические рули); разница "всего лишь" в совместном их действии в определенной комбинации и мере. Да и в нашем "типировании" к этому будут примешиваться другие факторы, в т.ч. обучение и инструктаж пилотов (аналог обучения/тренировки и сознательного выбора функций в соционике). Продолжая аналогию с психикой, пилот – это сознание, двигатель и органы управления самолетом – физиологическая основа, базисные механизмы реализации всех функций. Но здесь можно выделить еще один, промежуточный уровень – параметры системы управления (в т.ч. автопилота; пример – ограничения, введенные конструкторами и эксплуатантами с целью недопущения некоторых рискованных, но вполне возможных на данном самолете маневров). На этом уровне параметры определяют в какой мере и в каком качестве должны работать органы управления самолетом при маневрировании. К примеру, в учебно-боевом Як-130 изменением параметров системы управления можно сымитировать полет на других разнотипных истребителях.
Пример из другой области. Как соотносятся структуры компьютерных языков высокого уровня с архитектурой процессоров (а также – с машинным языком низкого уровня)? Опять же, связь есть, но не однозначная. Условные переходы, циклы, структуры данных реализуются определенными инструкциями процессора с определенным способом адресации памяти. Но соответствие "хард"- и "софт"-уровня (а также машинного и высокоуровневого языков) имеет и немало степеней свободы. Из одной и той же "высокоуровневой" программы можно получить разный машинный код, это зависит прежде всего от параметров (опций) компилятора (аналог параметров системы управления из предыдущего примера; от них зависит, к примеру, модель и способ адресации памяти).
Общее в этих примерах то, что соответствие "верхнего" и "нижнего" уровней опосредовано и может меняться (настраиваться) в определенных пределах. Верхний уровень, несомненно, использует структуры нижнего уровня в качестве базисных ресурсов. Без них он работать не может, они необходимы. Кроме того, ресурсы нижнего уровня специфичны, то есть ограничены в применении, их нельзя использовать по-всякому. Но! – специфичны они не полностью. Их можно использовать пусть и не произвольно, но в разной мере и в разном качестве. Последнее детерминируется промежуточным (передаточным) уровнем – параметрами системы управления (или компилятора).
Вернемся теперь к нашим баранам, то есть к феноменам психики и к соционическим функциям в частности. У разных систем и ресурсов мозга есть, в общем, базисное назначение в плане поведения, как правило, уже изученное нейропсихологией и когнитивной нейронаукой. К соционическим признакам и функциям их уже обстоятельно примерял В. Таланов [14, 16, 17, 18]. Но я считаю, что это именно базисные, самые "нижнеуровневые" ресурсы, и их описание – всего лишь половина решения задачи. Я полагаю, что между верхним уровнем (поведение) и нижним (специфические структуры мозга) существует промежуточный, передаточный уровень, на котором осуществляется интеграция и взаимная регуляция специфических систем и ресурсов нижнего уровня.
Более того, можно выступить с куда более сильной гипотезой, утверждая, что соционический тип личности определяется не базисными системами мозга (как считает Таланов), а именно промежуточным/передаточным уровнем интеграции базисных систем. Правда, это утверждение в большей степени зависит от того, что именно мы подразумеваем под ТИМ человека и, конкретнее, под информационным метаболизмом – т.е., от конвенции определения социотипа. К сожалению, данный вопрос в соционике пока либо замалчивается, либо забалтывается, но не решается конструктивно. Так что здесь придется аргументировать и этот выбор: что понятие информационного метаболизма можно (и даже нужно!) привязать к промежуточному уровню интеграции/регуляции функциональных систем (к самим этим системам можно неплохо привязать соционические функции, что делал и Таланов, и я в [7, 8, 10]).
В принципе, все подсистемы мозга работают параллельно, и соционические функции в своем базисном виде – не исключение. Причем ТИМ, вне зависимости от базовой функции, влияет тоже на все: на выражение эмоций, на стиль движений, на лексику речи, на сенсорное восприятие, на склонность к тому или иному типу мышления, – ни одна из функций не выключается, но все они модифицируются. Следовательно, уровень (внутреннего) информационного метаболизма – это надстройка над всеми функциями, модифицирующая параметры их работы и задающая порядок их взаимодействия (к примеру, описанные в [9] мотивационные связи и блоки), а не просто доминирование базовой функций над всеми остальными (вообще-то доминанта в мозгу переменчива и переключается по ходу ситуации).
Ранее я предполагал, что соционический тип определяется асимметрией важных межцентровых связей в мозге – сначала относил это на счет таламо- кортикальных связей [9], потом – на счет связей префронтальной коры с другими зонами [10, 11]. В принципе, асимметрия связей – достаточно общий (и, кстати, требующий дальнейшего уточнения) принцип реализации указанного промежуточного уровня регуляции. Ниже я опишу дейстие еще одной важнейшей интегрирующей структуры мозга, обеспечивающей связь префронтальной коры (здесь находятся "многомерные" исполнительные центры, см. [10, 11]) с задними "воспринимающими" отделами мозга. Это так называемое предклинье (англ. precuneus).
Отметим очень важную особенность указанных структур "передаточного" (в нашем представлении) уровня: все они эволюционно молоды, несоразмерно развиты у человека (и несколько меньше – у других высших приматов) по сравнению с прочими млекопитающими. Здесь входят: ассоциативный таламус, префронтальные исполнительные центры, теменно-височная область (угловая и супрамаргинальная извилины, примыкающая к ним зона Вернике, ответственная за речь), наконец, предклинье. Эти области относятся к третичной ассоциативной коре, не связаны с конкретными анализаторами и осуществляют высшую интеграцию информации из всех специфических областей. Они дают мозгу возможность не просто обрабатывать информацию, а делать это все более и более произвольным образом, в т.ч. перекладывать часть задач на функции, "изначально" имеющие совершенно другое назначение (в [7, 9] я показал это для блоков функций). Таким образом усиливаются потоки информации и активации между "старыми" специфическими системами мозга, задействованные в качестве уже мобильных (по задачам) ресурсов. В [9] я полагал, что именно этот обмен информацией/активацией стоит в основе понятия "информационный метаболизм".
В эволюционном плане развитие указанных структур, скорее всего, было продиктовано усилением социальности в эволюционном ряду приматов. Начиная с высших обезьян, социальность становится тотальной [21] сразу в двух смыслах. Во-первых, это постоянная включенность особей в общении (у прочих видов акты и периоды сигнальной коммуникации – конфликты за территорию, брачные игры – чередуются с периодами "асоциальности", когда особи взаимодействуют по минимуму). Во-вторых – почти все поведение становится потенциально сигнальным и социальным, в отличие от прочих видов, у которых есть четкое разделение на "социальные сигналы" и утилитарные действия – кормление, чистка, убегание и т.д. Когда я в [7] очерчивал этологический базис соционических функций, имел в виду как раз "старое" их назначение (в частности, социально-сигнальное назначение имеет базис рациональных функций, в отличие от "утилитарной" сенсорики). В своем базисе функции в целом самодостаточны и не особо нуждаются в интегрирующей надстройке, да и относительно простое поведение животных не требует такой интеграции: для каждой задачи есть своя программа действия, функции вполне могут работать обособленно, по принципу доминанты. В условиях же человеческой "тотальной социальности" все драматически меняется. К примеру, этическое отношение можно выражать не только стандартными эмоциональными выражениями, но вообще всеми функциями, самыми разнообразными средствами; само же отношение может распространяться не только на особей из своей группы, но на всевозможные объекты, в т.ч. неживые. То есть, этическая функция у человека – это не только ее базис (унаследованный у животных предков, пусть и сильно усовершенствованный), а сложный интерфейс взаимодействия базиса ("старой" этики) со всеми остальными функциями, позволяющий базисной этике использовать их возможности в своих целях; то же самое для остальных функций. Поэтому решающую роль здесь играет интегрирующий (передаточный) уровень, а не базис.
Наконец, тотальная социальность подразумевает тотальный же обмен сигналами между особями внутри группы, постоянный поток информации извне. Это – внешний, социальный информационный метаболизм, неизбежно порождающий свое отражение внутри психики – сложные модели типа "theory of mind". Вообще говоря, человеческий интелект развивается (по Л.С.Выготскому) от социального к индивидуальному; поэтому функции в их новом, интегрированном виде следует рассматривать прежде всего в социальном плане, как средства коммуникации и воздействия на поведение других особей, и лишь потом – как инструмент индивидуального мышления, оценки и восприятия ситуации. Иначе говоря, в соционике первичен социальный уровень личностного типа; интеллектуальный же уровень возникает, как приспособление "социальных" функций для решения несоциальных задач, физический урвень – вследствие вовлеченности тела в общении индивидов (первая сигнальная система). Правда, в соционике пока доминирует обратный подход – описание функций прежде всего в плане инструментального интеллекта и уже потом – социального действия. По нашему мнению, это является ошибкой (пусть не смертельной для соционики, но все-таки) и ее надо исправлять.
В нейронауке существует понятие default network (или default mode
network, default system [23]). Этим термином обозначается сеть корковых зон
мозга, активных в состоянии расслабленого бодрствования, когда человек не
сфокусирован на внешний мир и на решение внешних задач (иногда используют
термин task-negative network, TNN). Когда человек решает конкретную задачу,
эта система мозга деактивируется и активируется другая сеть (task-positive
network, TPN; эти две сети можно также рассматривать как компоненты единой
default network). В русском языке для этого термина пока нет устоявшевося
перевода; на мой взгляд, лучшим вариантом будет сеть фонового мышления,
или система фонового мышления, СФМ), но здесь буду использовать английское
сокращение DN.
На счет активности DN относят поддержание потока спонтанных мыслей в
сознании (в частности – пассивное воображение, мысли о будущем, воспоминания
прошлого), а также способность оценить и принять точку зрения другого
человека. DN отрицательно скоррелирована с системами мозга, занятыми
обработкой внешних сигналов, в особенности визуальных. Иными словами, ее
можно без особых натяжек отнести к базису соционической интуиции.
Отмечена также связь низкой активности DN с аутизмом, а высокой ее
активности – с шизофренией.
DN включает в себя участки префронтальной коры (ответственные за т.н.
theory of mind – моделирование психики других людей), часть медиальной
височной коры (память), задней цингулярной коры (эмоциональная оценка), а
также участки теменной коры вместе с т.наз. предклиньем (precuneus). На
последнее, в связи с его особой важностью, остановимся более подробно.
Предклинье – это участок теменной коры, спрятанный внутри борозды,
разделяющей оба полушария мозга (является частью поля 7 по Бродману).
Функционально оно состоит из трех отделов [24]. Передний отдел предклинья
выполняет сенсомоторные функции и связан с сенсорными, моторными и
премоторными зонами коры. Задний отдел связан со зрением и зрительной
корой. Но самые интересные функции выполняет центральный отдел. Он связан с
нижнетеменной корой (в частности, с угловой извилиной), с префронтальными
полями 8 (управление движением глаз), 10 (фронтополярная область), 46
(дорсолатеральная ПФК) и не имеет связей с сенсорными и моторными зонами.
За счет связи с дорсолатеральной ПФК центральный отдел предклинья вовлечен
в работе исполнительной системы мозга (в [10] я предположил, что эта
система ответственна за проявления т.наз. высших размерностей соционических
функций), а также в рабочей памяти и планировании движений.
Предклинье также имеет связи с подкорковыми структурами, в т.ч. с
дорсальными ядрами таламуса.
Исследования активности мозга в разных задачах и состояниях указывают
на весьма существенную, стержневую роль предклинья в процессах сознания,
самосознания (self-consciousness, self-awareness), рефлексии, в т.ч.
оценки взгляда на собственные поступки и качества со стороны других людей.
При инактивации предклинья человек теряет сознание. По современным данным,
предклинье составляет ядро описанной выше сети фонового мышления (DN), что
и объясняет важнейшую его роль для сознания. Считается, что предклинье
является важнейшим узлом связи (хабом) между префронтальными и теменно-
височными регионами коры. Этот хаб активируется многими когнитивными
функциями в разнообразных задачах [23, 24].
Особо отметим связи центрального участка предклинья (на рис.1
обозначены зеленым). В сторону лобных долей они ведут к дорсолатеральной
префронтальной коре, ответственной за исполнительные функции ("усилитель
мышления", реализующий высшие мерности функций с т.з. соционики). Но также
есть связь с нижнетеменной корой, – а это важнейший участок, ответственный
за речь (не только за смысл, но и за строение речи), за восприятие метафор,
за математические способности, за восприятие мотивации других людей (theory
of mind), отчасти – за взгляд на собственное тело со стороны (эксперименты
с "выходом души из тела" при стимуляции угловой извилины). Ранее В. Таланов
связывал левую (речевую) область теменно-височной коры с интуицией
[14]; опять же с интуицией он связал и правую лобную кору [19],
что, в общем, сходится с перечисленными выше фактами.
Однако прочие отделы предклинья поддерживают связи и с сенсорной/
моторной, и с лимбической (цингулярной) корой, ответственной за эмоции, см.
рис.1. Кроме того, еще в [9] я отмечал, что поток свободных мыслей в
голове не обязательно интуитивен в аспектном смысле – ведь мысли могут быть
очень даже сенсорного содержания. Аналогично, хотя языковой знак "сам по
себе" интуитивен (в смысле отсутствия сенсорного сходства означаемого с
означающим), его означаемое (= смысл речи) может входит в любой аспект. Так
что скорее всего здесь мы сталкиваемся со структурой двойного назначения:
нижнетеменние доли, "изначально" нацеленные на решение интуитивных задач,
по ходу развития связей центрального отдела предклинья и углубления
интеграции с остальными функциями мозга, приобрели возможность обрабатывать
информацию по любому аспекту. Аналогично, дорсолатеральная ПФК изначально
входила, скорее всего, целиком в сферу логики, но впоследствии, по ходу
углубления интеграции мозговых систем, получила возможность "усиливать"
мышление любой другой функции.
В итоге вся система "центральное предклинье – нижнетеменная кора –
дорсолатеральная ПФК" стала настраиваемой. Ее параметрами, согласно
нашему подходу, являются связи ПФК и НТК со специфическими отделами
коры, где хранится не только информация по каждому аспекту, но также смысл
слов, входящих в этот аспект. (Известно, что словарный запас разных
словоформ в основном "разнесен" по разным участкам коры; так, смысл
глаголов ( Еще раз перечислим основные структуры, входящие в DN и определяющие
характеристики спонтанно-фонового мышления:
Будем называть их структурами внутреннего информационного
метаболизма психики.
Почему я выбрал такое название, а не считаю эту систему просто
интуитивной (см. выше)? В [9] я вводил понятие интенсивности
информационного метаболизма, как условная мера "количества мыслей в
голове", и еще тогда отмечал, что оно соотносится с интуицией в понимании
немалой части социоников, в частности, Таланова. Но кроме количества есть и
качественные характеристики потока мыслей – преобладание мыслительной
активности по одним аспектам над другими (в т.ч., преобладание этих
аспектов во внутренней речи, да и во внешней). Большая общая "интенсивность
мыслей" отнюдь не подразумевает, что последние сдвинуты к интуитивным
аспектам; они могут быть и вполне сенсорными, с задействованными высшими
мерностями сенсорики, а не интуиции, с активационным блоком по сенсорике и
проч.
Ранее в [11] я анализировал два типа факторов, влияющих на развитие
межцентровых связей в мозге (и оттуда, на формирование ТИМ) – генетические
и эпигенетические, в частности, материнский эффект. Суть последнего можно
увидеть на статистике Е.С. Филатовой [20], где, кроме большого количества
детей, по ТИМу тождественных отцу (27%, что говорит о несомненном
генетическом вкладе в ТИМ), было достаточно много детей, дуальных либо
противоположных матери (33%). Этот эффект зависит от пола ребенка и лучше
выражен у сыновей (41%); у дочерей он несколько видоизменен – дуалов и
погашенцев матери встречаются чуть реже, но зато есть более общий обратный
эффект для признака экстра-/интроверсии – в 2/3 из случаев вертность дочери
противоположна вертности матери.
Если предположить, что этот материнский эффект (МЭ) закладывается не
внутриутробно, а в самом раннем постнатальном периоде (первые недели жизни
ребенка), его можно рассматривать как социальную адаптацию: мозг ребенка в
некоторой мере подстраивается под стиль общения матери, что может улучшить
их взаимодействие. Специально оговоримся – это не воспитание и не
подражание, ведь это первые недели жизни, да и ребенок не копирует мать, а
наоборот, несколько отдаляется от нее по стилю общения (в частности,
тяготеет к противоположной вертности). Скорее всего, сила МЭ может меняться
от случая к случаю, т.к. поведение и стиль общения матери зависит отнюдь не
только от ее ТИМ, но и от ситуативных и культурных факторов.
В [11] я предложил примерную схему формирования ТИМ как
произведения двух отдельных групп факторов: темперамент (вертность +
нальность) и установка (клуб). Предполагалось, что МЭ влияет и на те и на
другие, но в разной форме. Однако углубленный анализ реальной статистики
Филатовой выявил гораздо более сложную картину. Наименее подверженными МЭ
оказались признаки Рейнина рацио-/иррациональность и демо-/аристократия (и
дополнительный им признак процесс/результат). МЭ, кроме вертности, сильно
влияет на ПР квестимность/деклатимность, но только у сыновей. На признак
Юнга сенсорика/интуиция МЭ влияет вместе с генным фактором, а логика/этика
определяется, судя по всему, демо-/аристократией и сформированной уже
сенсорикой/интуицией (хотя детали этого взаимодействия выяснить до конца не
удалось). Детальный анализ данных Филатовой представлен в
приложении.
Согласно данным нейрофизиологии и нейрогенетики, в развитии разных
структур мозга в разной мере участвуют генные и средовые (к которым по сути
относится и МЭ) факторы. Генными факторами в наибольшей мере определяется
развитие подкорковых структур; средовыми – близкие связи (между смежными
зонами коры и внутри одной и той же зоны); в развитии дальних связей (между
отдаленными участками коры, а также от коры к подкорке) сопоставимый вклад
вносят факторы обеих групп [4]. В рамках гипотезы, что ТИМ задается
балансом связей default network, и зависит как от МЭ, так и от
генетики, логично заключить, что он должен определяться дальними связями.
Но анализ лучше начать с признаков, наименее зависящих от МЭ – нальность и
кратия. Для первого из них В. Таланов показал, что рациональность связана с
преобладанием активности передних отделов мозга, иррациональность
(воспринимающие функции) – с задними и височными [17]. Вопрос: с какими
структурами мозга можно связать второй признак – кратию (или дополняющий ее
ПР процесс/результат)? Точнее: не является ли один из этих признаков
отражением другой фундаментальной асимметрии мозга – лево-правой?
Эта идея родилась из того, что в [10, 11] я уже рассматривал
модель, состоящую из 4 отдельных исполнительных центра мозга – в левом и
правом полушарии, в префронтальной и нижнетеменно-височной областей. Таким
же образом можно выделить и четыре мотивационных центра (МЦ), функция
которых – интеграция и координация работы коры (в т.ч., фрагментов системы
фонового мышления) в соответствии с конкретными задачами и с мотивацией
человека. Два из них находятся в задней цингулярной коре (слева и справа), об
их действии в составе DN см. [23]. Остальные два будут либо в
передней цингулярной коре (anterior cingular cortex, поля по Бродману 24, 32, 33,
см. рис.2), либо, как вариант, в вентромедиальной префронтальной коре (часть
поля 10), тоже симметрично в двух полушариях. Рассмотрим по отдельности
особенности работы передних и задних отделов полушарий, описанные в
литературе [1, 13].
Эксперименты показывают, что лобные доли в наибольшей степени активны
при решении новых задач, либо сложных задач, требующих особой концентрации.
По мере привыкания и рутинизации активность смещается к задним, менее
мощным и оттого менее энергозатратным центрам [1]. Из-за этого на активность
лобных долей долями можно отнести многомерность мышления (в соционическом
смысле). Однако многомерность обеспечивается не мотивационными структурами,
а префронтальными исполнительными центрами (поля 9, 46; см. выше, а также
[22]). Последние через систему DN связаны и с задними МЦ, то есть
многомерность не является эксклюзивным свойством передних МЦ.
Также считается доказанным, что активность лобных долей связана с
волевым усилием, с концентрацией внимания и воли на конкретную решаемую
задачу [1]. То есть, задние мотивационные центры более спонтанны в своей
работе, передние – более планомерны, что в соционике отражается в признак
рацио-/иррациональность (как давно заметил В. Таланов). Можно предположить,
что у рационалов активнее передние МЦ, у иррационалов – задние, но и те и
другие могут иметь доступ к высшим мерностям функций.
Лево-правая асимметрия активации традиционно связывается с речевым/
неречевым либо сознательным/неосознанным мышлением (у "стандартных
правшей"). Однако есть и другие особенности. Согласно [1], правые лобные
доли активируются в ситуации новизны и, что важнее, неопределенности:
когда априори неизвестно какой именно цели добиваться, какими критериями
успеха придерживаться, какими средствами пользоваться. Левые же лобные доли
активируются при решении пусть и сложных либо новых задач, но по образцу
– когда извне задана цель, критерий оценки и/или способ решения. (Позже, по
мере рутинизации, активность мозга в таких задачах смещается к задне-правому
центру, запускающему уже выработанный автоматизм в нужной ситуации [13]).
Но если способ решения задачи априори не указан либо сама задача поставлена
в свободной форме, включается правый лобный центр вместе с левым задним –
диагональ "творческого мышления" [6]. Левый задний центр, скорее всего,
координирует решение задач в уже устоявшейся ситуации и знакомыми
инструментами, правый передний осуществляет выбор целей, критериев и
инструментов решения в новых ситуациях.
Согласно [1], лево- и правополушарная стратегии принятия решений
(т.наз. когнитивные стили) отличаются своим адаптационным назначением. Суть
левополушарной (по [1] – контекст-зависимая) стратегии – в стремлении
как можно лучше "подогнать" принимаемые решения под конкретные особенности
текущей ситуации (= контекста); оттуда и стремление учесть как можно больше
подробностей. Эта стратегия на первый взгляд кажется единственно
оптимальной; однако она перестает работать, если ситуация радикально
меняется и старие критерии оценки, вместе с наработанным опытом, перестают
работать. В таких услових уже неизвестно какие методы правильны, какие
критерии оценки надо использовать. И тут продуктивнее оказывается
правополушарная (контекст-независимая) стратегия: игнорирование деталей
ситуации, следование более общим критериям, потенциально применимым в
широком диапазоне условий. Цель – относительно быстрая реакция в незнакомой
ситуации, без "зависания" и долгих обдумываний.
Несложно увидеть сходство этого описания лево- и правополушарной
стратегий с соционическим признаком "левые/правые", в семантику которого
входит как раз стремление к упрощению либо усложнению [2]. При этом
левополушарная стратегия оказывается соционически "правой", правополушарная
– "левой". Можно увидеть и еще одно проявление того же признака Рейнина –
процесс/результат. Левополушарная стратегия подразумевает ориентацию на
процесс – обдумывание пошаговых планов либо действие в череде ситуаций,
меняющихся лишь в заданных пределах, что позволяет быстро учитывать каждое
изменение ключевых факторов. Правополушарная стратегия, наоборот, состоит в
отказе от втягивания в процесс, в прерывании лишнего обдумывания и в
принятии полученного результата именно как результат (а не как исходный
пункт очередного раунда обдумываний, чем часто грешат "правые" ТИМы).
В соционической литературе возможность привязки признака "процесс-
результат" к полушариям мозга упомянута В. Мироновым [5], но без ссылки на
конкретного автора идеи, лишь в виде упоминания о существовании такой
гипотезы на раннем этапе развития соционики.
В принципе, активность левого и правого полушарий может чередоваться и
переключаться по ходу ситуации: в знакомых условиях доминанта активности
смещается влево, в незнакомых либо неожиданно меняющихся – вправо. Конечно,
у разных людей относительная активность полушарий и мотивационных центров
(в лево-правом и передне-заднем направлении) может отличаться – это и есть
одна из форм межиндивидуальных отличий [13]. Она определяет способности
мозга работать в разных режимах и предпочтения разных когнитивных стилей
[1]. Так, высокая активность левой лобной коры считается оптимальной для
деятельности, требующей высокой концентрации внимания и выбор правильной
реакции в сложной, но знакомой обстановке (операторская деятельность). Если
активнее задний центр мотивации, это проявится в склонности к упрощению и
автоматизации реакций. Высокая активность правой лобной коры, скорее всего,
выливается в склонность к "творческому" решению задач по своему усмотрению
(в отличии от активности левого лобного центра, работающего в основном по
узким, заданным извне критериям).
Здесь надо сделать очень существенное уточнение, касающееся активности
четырех МЦ. Она, разумеется, может меняться по ситуации. Однако нам здесь
интересны не МЦ и не полушария сами по себе, а их роль во внутреннем
информационном метаболизме. Роль эта определяется связями МЦ с сетью DN
(в т.ч., с предклиньем), и эти связи относительно устойчивы и малоизменчивы
(по сравнению с активностью соединяемых участков, которая меняется очень
широко по ходу ситуации). То есть, в период доминирования активности сети
DN (= режим фонового мышления) будет задействован тот МЦ, который связан с
DN сильнее прочих. Как следствие, в работе DN проявятся соответствующие
полюса рацио-/иррациональности и демо-/аристократии. Связи МЦ с DN, будучи
дальними (см. выше), должны определяться в основном генными факторами, с
влиянием материнского эффекта в некоторых случаях (о наследовании
нальности и кратии, см. приложение).
От признаков нальности и кратии зависит вид мотивационных блоков –
связей, обеспечивающих автоматическое включение базовой функции в ответ на
активность творческой. С учетом сказанного о локализации признаков по
полушариям получается следующая схема локализации мотивационных блоков по
мотивационным центрам:
К примеру, если ведущим является левый передний МЦ, сенсорное восприятие
вещей автоматически будет включать логику, причем логическая оценка и
анализ объектов (а также составление планов действия) станет основной
задачей, оттеснив на второй план собственно восприятие. Но если ведущим
является правый передний МЦ, сенсорные сигналы будут трансформироваться
прежде всего в эмоциональную оценку, а логика будет включаться в ответ на
интуитивные сигналы-ассоциации. Передние МЦ включают рациональные,
оценочные функции в ответ на сенсорное восприятие и на интуитивные
ассоциации; задние же "иррациональные" МЦ, наоборот, включают сенсорику
(действия, поисковая активность) либо интуицию (ассоциативный аппарат) в
ответ на логические или эмоциональные сигналы.
Отметим, что четыре блока модели А – Эго, Ид, Суперэго, Суперид –
оказались "втиснутыми" в один мотивационный центр. Эта на первый взгляд
второстепенная деталь может оказаться ключевой для обоснования модели А с
точки зрения нейрофизиологии. Точнее: данная четверка мотивационных блоков,
возможно, способна работать одновременно без переключения МЦ и, стало
быть, без "обрывания" режима DN, к которому привязан ведущий МЦ. Конечно,
не все четыре возможных блока внутри данного МЦ смогут работать одинаково,
в одном и том же качестве: один из них будет доминировать, одна пара
функций окажется многомерной, другая – маломерной.
Выбор ведущего блока внутри МЦ, судя по данным Филатовой, определяется
при рещающем участии материнского эффекта, но и с влиянием генных факторов
(см. приложение). Наиболее вероятный механизм дифференцирования блоков –
выделение внутри МЦ оппозиции "воспринимающего" и "действующего" блоков:
первый отражает действия и сигналы матери, второй управляет активностью
самого ребенка и его сигналами, посредством которых он общается с матерью.
"Действующий" блок (Эго) чаще всего устанавливается в Суперид-отношение с
воспринимающим, если судить по преобладании дуального отношения ребенок-
мать. Таким образом МЭ "доопределяет" оставшиеся признаки ТИМ: вертность и
один из пары логика/этика и сенсорика/интуиция (второй дозадается демо-/
аристократией). Но это не единственно возможное отношение "действующего" и
"воспринимающего" блоков. Так, они могут установиться в Ид-отношении
(противоположность ребенок-мать – тоже частый случай в данных Филатовой). К
тому же "действующий" и "воспринимающий" блоки могут оказаться вообще в
разных МЦ – особенно в случае гетерозиготности по фактору, кодирующему
мощность связей МЦ с DN. Тогда и отношения ребенок-мать будут другими, и,
что гораздо важнее, – оставшиеся признаки ТИМ могут определиться не МЭ, а
генными факторами. (Доказательство тому – частая тождественность ребенка
отцу; кроме того, на данных Филатовой МЭ по вертности "работает" только при
одинаковой нальности матери и ребенка. Ограниченный объем статистики не
позволил мне разобраться до конца с закономерностями наследования всех
признаков ТИМ.)
Разнообразие возможностей для отношения воспринимающий-действующий
блок в период действия МЭ (а это, предположительно, первые недели жизни
ребенка) поднимает ряд серьезных вопросов, выходящих за рамки существующих
соционических теорий. К примеру, может оказаться, что характер работы
функций и блоков зависит не только от их положения по отношению к ведущему
блоку (собственно то, что описывается моделью А), а еще от взаимодействия
конкретных факторов (генных и МЭ), сформировавших настройки информационного
метаболизма. Можно рассматривать это и как подтипную вариативность, и как
источник возможных нестыковок реального взаимодействия функций с моделью А.
Эти вопросы крайне интересны, но на сегодняшнем уровне соционики очень
сложно не то что решить, но даже сформулировать их в достаточно
конструктивном виде; тому мешает ограниченность понятийного аппарата,
нацеленного на описании лишь признаков из базисов Юнга и Рейнина, но не
механизмов их работы и формирования.
Еще одна возможная паралель взаимодействия МЦ с моделью А:
периодическое чередование активности двух "однополушарных" МЦ может
трактоваться, как чередование фаз информационного метаболизма по ревизному
или заказному кольцу. (Замечание: Я не считаю, что сеть DN намертво
привязана только к одному МЦ, а остальные ей полностью недоступны. Более
естественным кажется допущение, что DN поддерживает связи со всем МЦ и
ситуационно может их активировать. Но относительная слабость связей ведет к
тому, что эта активность не может продолжаться достаточно долго, в конечном
итоге "скатываясь" к "главному" МЦ. Но если связи двух МЦ с DN сравнимы по
мощности, это может выражаться в периодическом переключении двух режимов,
как в описанном случае с кольцами заказа/ревизии. Если же два сильнейших МЦ
находятся по диагонали друг к другу, это приведет к чередованию рацио- и
иррациональных режимов одной и той же кратии: к примеру, зеркальных пар
Наконец, можно рассмотреть в качестве отдельного подтипного фактора
активность центрального узла связи сети DN – предклинья. Чем он активнее,
тем сильнее обмен информацией между удаленными участками коры, – вне
зависимости от асимметрии самих связей DN с МЦ (а также с зонами, где
хранится информация по отдельным аспектам, с центрами активации и тд). Это
в очень большой степени похоже на признак сенсорика-интуиция, но я уже
говорил, что на уровне теории активность мозговых систем надо относить к
подтипу, а к собственно ТИМу – только баланс их связей [9]. То есть
активность DN в целом определяет подтипную интуицию/сенсорику –
"количество мыслей в голове", по выражению В.Таланова, в свою очередь очень
близкую к дихотомии информация-энергия у В.Гуленко [3]. Возможно,
это определяется генным фактором (см. приложение).
В заключение надо сказать, что описанная схема мотивационных блоков
и их распределение по полушариям мозга пока является гипотетической. Она
следует главным образом из статистики Филатовой, в которой одни признаки
(нальность, кратия и "замыкающий" их процесс/результат) показали более
"классические" (с т.з. генетики) закономерности наследования, тогда как на
остальные признаки очень сильно влияет материнский эффект, являющийся по
сути влиянием среды, а не собственно генным фактором. По ходу обдумывания
этих вопросов я останавливался и на других гипотезах. Сначала полагал, что
ТИМ определяется настройками DN только речевого (в норме, левого) и более
"сознательного" полушария, поскольку они могут задавать как предпочтения в
структуре речи по аспектам, так и большую либо меньшую "осознанность" тех
же аспектов. Потом рассматривал возможность распределения четырех блоков
модели А по четырем МЦ, причем блок Эго оказался слева/впереди, Ид – справа
/впереди, Суперид – слева/сзади, Суперэго – справа/сзади (многомерные блоки
– впереди, маломерные – сзади, ценностные функции, они же вербальные, –
слева, неценностные – справа). В этих случаях материнский эффект задавал бы
настройки левого заднего центра (предположительно Суперид, в согласии с
высокой вероятностью формирования психики, дуальной материнской). Потом
рассматривал разные схемы распределения блоков по МЦ (не только по модели
А), причем оно должно было зависеть от генов, полученных от матери и отца,
а также от МЭ, и в теории возникало что-то вроде внутритипного отношения
между генными и "установочными" (МЭ) факторами формирования ТИМа. Но позже
я отказался от этих схем в пользу фиксированного распределения блоков по
МЦ. (Еще раз отмечу, что лево- и правополушарность МЦ совершенно не связана
с речевыми функциями или с дуализмом "сознание – бессознательное". Речевые
функции – это конкретные зоны коры (Брока и Вернике), активность которых
односторонне слева коррелирует с активностью центрального предклинья (см.
рис.1 выше), тогда как МЦ располагаются в лимбической коре, не связанной
напрямую ни с речью, ни с "сознанием".) Здесь я решил поделиться этими
рабочими гипотезами, т.к. другим соционикам они могут показаться более
правдоподобными, да и сам вопрос ни в коем случае не может считаться
окончательно решенным.
июль-декабрь 2012
Статьи из Википедии:
Сеть фонового мышления
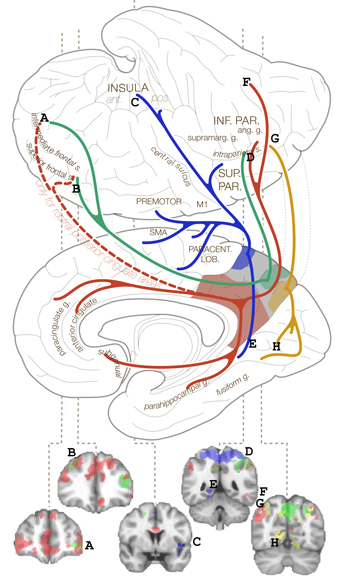
Фиг.1. Связи разных отделов предклинья с другими зонами коры
мозга (схема взята из [24]).  ) в основном хранится в левой лобной коре, существительных
(
) в основном хранится в левой лобной коре, существительных
( ,
,  ) – в правой среднетеменной коре, предлогов (
) – в правой среднетеменной коре, предлогов ( )
– в левой теменной коре.) Эти связи в итоге задают предпочтение и приоритет
одних аспектов перед другими: настройки ПФК определяют размерность функций,
настройки НТК – предпочтения в употреблении аспектной лексики. Возможно,
настройки эти на самом деле едины и определяются интегрирующим узлом сети
DN – предклиньем. (К примеру, путем асимметричного замыкания центрального
отдела предклинья на связи остальных отделов со специфическими моторными,
сенсорными и эмоциональными зонами; здесь также возможно участие таламуса,
своей активацией "подсвечивающего" нужные участки коры по "указаниям
сверху", из предклинья.) Но теоретически возможен и случай (хотя бы)
частичной независимости пораметров настройки ПФК и НТК, когда многомерными
бывают одни функции, а лексика тяготеет к другим аспектам.
)
– в левой теменной коре.) Эти связи в итоге задают предпочтение и приоритет
одних аспектов перед другими: настройки ПФК определяют размерность функций,
настройки НТК – предпочтения в употреблении аспектной лексики. Возможно,
настройки эти на самом деле едины и определяются интегрирующим узлом сети
DN – предклиньем. (К примеру, путем асимметричного замыкания центрального
отдела предклинья на связи остальных отделов со специфическими моторными,
сенсорными и эмоциональными зонами; здесь также возможно участие таламуса,
своей активацией "подсвечивающего" нужные участки коры по "указаниям
сверху", из предклинья.) Но теоретически возможен и случай (хотя бы)
частичной независимости пораметров настройки ПФК и НТК, когда многомерными
бывают одни функции, а лексика тяготеет к другим аспектам.
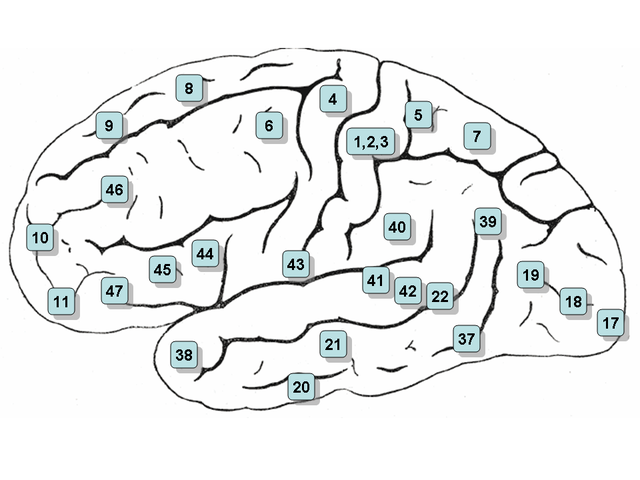
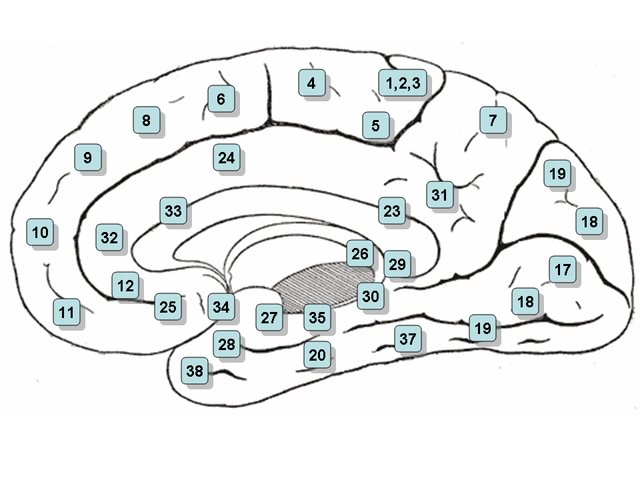
Фиг.2. Карта латеральной (наружной) и медиальной (внутренней)
поверхности коры мозга с обозначением полей по Бродману (схема из Википедии,
Brodmann area).
Мотивационные центры и их локализация по полушариям мозга
Мотивационные центры и модель "А"
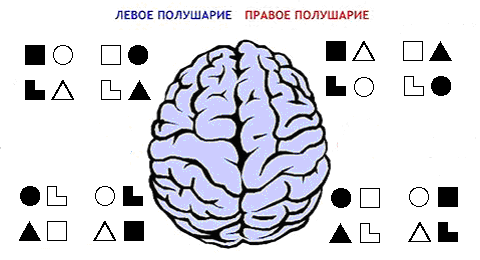
Рис.3. Распределение мотивационных блоков по полушариям мозга.
– для
"аристократической" заднеправо-переднелевой диагонали, или
 –
для "демократической" заднелево-переднеправой диагонали, что сходится
с описанной в [6] картины активности диагоналей коры при "предметном"
и "творческом" мышлении. Сравнимая активность симметричных МЦ в двух
полушариях, наверное, приведет к чередованию активности "деловых" либо
"родственных" режимов мышления.)
–
для "демократической" заднелево-переднеправой диагонали, что сходится
с описанной в [6] картины активности диагоналей коры при "предметном"
и "творческом" мышлении. Сравнимая активность симметричных МЦ в двух
полушариях, наверное, приведет к чередованию активности "деловых" либо
"родственных" режимов мышления.)
* * *
Литература
2. Гуленко В.В.
Формы мышления (2002)
3. Гуленко В.В.
Энергетическая модель соционического объекта. Структура и функционирование. (2007)
4. Ивонин А.А., Цицерошин М.Н., Погосян А.А.,.Шуваев В.Т. Генетическая
обусловленность нейрофизиологических механизмов корково-подкорковой
интеграции биоэлектрической активности мозга. // Рос.физиол. журн.2002.
Т.88,№10. С.1330-1342. (тезисы здесь)
5. Кочубеева Л.А., Миронов В.В., Стоялова М.Л. Соционика. Семантика информационных аспектов. – СПб.: Астер Х, 2006. – 146 с.
6. Павлова Л.П., Романенко А.Ф. Системный подход к психофизиологическому
исследованию мозга человека. -Л.:"Наука" 1988.
7. Попов И.
Базис соционических функций и аспектов с точки зрения этологии и эволюционной
психологии (2009).
8. Попов И.
Позитивизм/негативизм, квестимность/деклатимность и функциональная асимметрия
мозга (2010).
9. Попов И. Связи (блоки)
соционических функций (2010).
10. Попов И.
Нейрофизиологическая природа размерности функций (2011).
11. Попов И.
Факторы формирования соционического типа личности и их взаимодействие (2011).
12. Равич-Щербо И.В., Т.М. Марютина, Е.Л. Григоренко. Психогенетика. //
Под редакцией И.В. Равич-Щербо. – М., 1999 //
с. 349-362.
13. Русалова М.Н.
Функциональная асимметрия мозга: эмоции и активация // Функциональная
межполушарная асимметрия (ред. Н.Н. Боголепова, В.Ф. Фокина). - М: Научный
мир, 2004.
14. Таланов В.Л. Новые представления о физиологическом механизме и локализации
базисных юнговских функций. // Соционика, ментология и психология личности,
2002, № 4.
15. Таланов В.Л.
Краткое изложение модели «Т»: физиологической модели информационного
метаболизма в психике человека
16. Таланов В.Л.
Структура признака «интуиты-сенсорики» и его гипотетические когнитивные
механизмы (2007)
17. Таланов В.Л.
Подробная кластерная структура, когнитивные и физиологические механизмы
признака "иррационалы-рационалы". (2007)
18. Таланов В.Л.
Маркеры доминирования эрготропной симпатической системы и нейромедиаторной
активности в проекции на соционические признаки. (2007)
19. Таланов В.Л.
Всё неизвестное и малоизвестное о восьми функциях психики. Часть I:
Расчёт функций, количественное значение всех функций в психотипе, содержательное наполнение функций. (2012)
20. Филатова Е.С.
Соционическая статистика для 299 женщин, мужчин и их детей //
«Соционика, ментология и психология личности», Киев, 2000, № 6 (33).
21. Фридман В.С.
«Человек» и «животное»: где провести грань?-1 (2010)
22.
Executive functions
23.
Default network
24.
Precuneus
Иван Попов, 2012
![]()